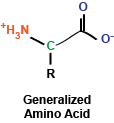In a previous post, we had explored the characteristics of
the 20 amino acids that make up proteins.
These amino acids make up a diverse collection of molecules that can be
strung together, making up proteins that have a multitude of functions within
the cell.
 Amino acids form a protein through the action of the
ribosome, which we will discuss in a future post. At this time, suffice it to say that the
ribosome uses an elegant mechanism to read mRNA and translate it into the
protein encoded by the mRNA by adding amino acids in a string. As this string of amino acids is created, it
begins to form a structure that will have functions within the cell.
Amino acids form a protein through the action of the
ribosome, which we will discuss in a future post. At this time, suffice it to say that the
ribosome uses an elegant mechanism to read mRNA and translate it into the
protein encoded by the mRNA by adding amino acids in a string. As this string of amino acids is created, it
begins to form a structure that will have functions within the cell.
Four levels of protein structure exist:
Primary (1o)
structure: The order of the amino
acids is the primary structure. Think of
the primary structure as the alphabet of the amino acids: MGRYNVPL, for
example. The primary structure describes
what order amino acids are in, and even though it might not seem like very much
information, even the primary structure of a protein can provide a great deal
of information in terms of its function and its potential 3-dimensional shape.
Secondary (2o)
structure: When amino acids are
polymerized, they form local structures, which make up the secondary
structure. Think of secondary structure
as the shape of a group of amino acids.
Two primary forms of secondary structure exist: alpha helices and beta sheets. Alpha helices result from the coiling of the
amino acid string turning about itself.
In contrast, beta sheets are flatter and lack coiling. There are several types of alpha helices and
beta sheets, which we will save for a future post, and these different types of
structures have important implications for overall shape of a protein.
Tertiary (3o)
structure: Protein structure gets
exciting when you talk about tertiary structure, which can be described as the
overall three-dimensional shape of a protein.
In general, the tertiary structure is the “final” form of a protein,
although modifications on the protein, as well as interactions with other
proteins can affect this structure.
Quaternary (4o)
structure: When proteins interact
with each other, they can form complexes, which is the quaternary structure of
these proteins. You can think of this
structure as the way proteins contact each other. The bundling of proteins together can be
between proteins of the same type (such as is the case with hemoglobin) or
other types of proteins.
The drawing attempts to illustrate the concept of the
different levels of protein structure.
Again, think of the primary structure as the order; the secondary structure
as the local shape; the tertiary structure as the overall shape; and the
quaternary structure as the way this tertiary structure of the protein
interacts with other proteins.
How protein structure is established is a fascinating
question and a field that is actively studied by prominent labs around the
world. Protein folding is the process of a chain of amino acids curling
into its final shape, and how this process occurs is complex and not completely
understood. In general proteins fold
depending on their environment (exposed to water or not, for example) and with
the help of other proteins, called chaperones. Protein chaperones help to establish a
protein’s structure as well as maintain it during times of stress. Further, modifications on proteins can change
their structures, such as when p53, a protein that is involved in regulating
many processes within the cell, is phosphorylated – its structure and,
consequently, its function is altered slightly.
The structure of proteins has fascinated scientists since we
first learned about proteins. Thousands
of scientists around the world are still working to discover new structures and
to learn how proteins fold. The science
behind protein folding has important implications in diseases such as Alzheimer’s,
cancers, and infectious diseases.